|
|
|
|
|
|
ウシの目の解剖
作業時間:約1時間 ただし、狂牛病問題のため事実上打ち切りの実験です!

■ はじめに ■
- 代表的な実験の一つです。
- 眼は、詳しく調べれば調べるほど奥の深い器官であることを実感します。是非生徒にも体験してもらいたいものです。
- 牛の目は他の動物よりも大きく、また、比較的入手が容易です。 そのため、解剖を行うには格好の素材です。
- 牛の目は瞳孔が四角形で、黄斑がないことを除き、ほぼヒトの目と類似しています。
- ただし、現在は狂牛病の問題があるため、教育現場でのこの実験は行われません。
■ 用意するもの ■
- ウシの目・・・食肉センターから入手しました。

- 解剖セット・・・写真の通りです。
- 新聞紙・・・テーブルが汚れないようにするためと、レンズで文字を見るときのため。
- ゴム手袋・・・あった方が良いですね。
- 消毒用アルコール・・・念のため。
■ 方法 ■
- 実験の展開にはいろいろな方法がありますが、今回は実験を5つぐらいのステージに分けて、ステージごとに指示しながら展開しました。
①余計な肉や脂肪の除去し、目をクリーニング。これを行って慣れさせます。
②角膜側と視神経側に、半分に切断。
③視神経側の観察。
④レンズの観察。
⑤角膜や毛様体の観察。
■ 結果 ■
- 牛の目玉を後ろから見たものなのですが、何十個もの目を同じ袋に入れたせいで変形しちゃってます。眼球自体は問題ありませんが。
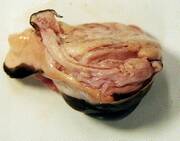
- 余分な肉をそぎ取ります。特に視神経のまわりをきれいにそぎ取り、視神経の伸び方を確認します。
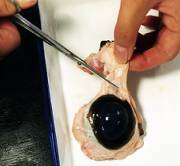
- 強膜をはさみで切ります。強膜は意外と強固なので、ピンセットで押さえるのは難しいです。この段階で、意を決して素手で抑える生徒もでてきます。
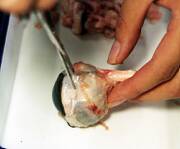
- 角膜側と視神経側に分かれるように切断すると、ゼリー状の硝子体がでてきます。
下の写真中の左側の視神経からつながっている、どろどろした肌色のものは網膜です。

- 中央が水晶体(レンズ)です。そのまわりにはチン小帯が張り付いています。
この目玉は授業で余ったもので、5日ぐらい経っているので、レンズが白く濁っています。(日数が経過するとレンズは白く濁ります。)

- 写真の中の上の方に視神経が伸びています。網膜の後ろにはきれいなエメラルドのようなテカリのある膜があります。ネコやイヌなどにはタペタムというものがあり、写真の通り、ウシにもあります。写真などを撮影したとき、ネコの目が黄色っぽく写ったり、イヌが緑っぽく写るのは、タペタムに光が当たり反射したときの色です。ヒトはタペタムを持っておらず、血管の色が写し出され“赤目”になります。

- タペタムは、網膜色素上皮の外層のブルーフ膜(網膜最外層)と脈絡膜最内層の脈絡毛細血管板の間にあります。すなわち網膜と脈絡膜の間ということです。

- タペタムと脈絡膜を除くと色素層(真っ黒)になります。ここが強膜と脈絡膜の境界です。
シャーレの左側にあるものは網膜ですが、ちょっと注意が必要です。解剖して硝子体を取り出すときに眼底から一緒に剥がれてくる半透明の薄い膜が網膜ですが、網膜は約8層の構造で、解剖のときは最外層の網膜色素上皮が強膜側に残ってしまいます。そのため正確に網膜,脈絡膜に分けることはできません。ただし、大学のような専門ではなく、目の解剖の基礎実験ですから、大雑把に説明をすれば良いと、私は思います。

- 新聞紙の上に水晶体(レンズ)を置くと、文字が大きくなって見えます。残念ながらこれは濁っています。

- 角膜側の断面を眼球内部から撮った写真です。
牛の目は瞳が長方形になっています。瞳孔を大きくしたり、小さくしたりして虹彩の働きを説明します。

- 視神経を縦に切断した写真です。
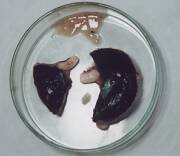
- 虹彩を除き、まっぷたつに切った写真です。

- 角膜の断面です。

■ コメント ■
- 生徒は、この解剖を行う直前に目の構造名をプリントに記入し、復習しました。 しかし、いざ解剖となると各部を判別するのが容易ではないようです。説明を加えると「あー、これか!」というような感じでした。
- 生徒の感想としては”気持ち悪かった”というのがたくさんありました。 しかし、以前にドッグフードのニワトリの脳を解剖していたし、血が出ないこともあり、私が考えていたよりも積極的に作業していました。
- タペタムを見たり、レンズで文字を見たり、角膜に接していた虹彩が剥がれるのを見たりすると、感嘆の声があがります。生徒はイラストや模式図で目の構造を学んでいましたが、色や形を自分の中でイメージしているものの、やはり実物の観察は有意義であったようです。
- 牛の目の解剖の場合、各部の構造を観察するのが目的です。2時間連続の授業で構造の復習からじっくりと取り組めたので、学んだ構造をほとんど確認できのではないかと思います。中には、レンズを取り出すのが目的と勘違いしている生徒がいたのは残念でしたが...
- この実験を行っていたのは1998年頃です。あれから数年経つと、狂牛病の問題が発生しました。現在、牛の目を教育現場で扱うのは相応しくないということで、おそらくどの学校でも行っていないでしょう。というよりも県教委からやらないようにとの指示がありますのでできません。もちろん、その代わりとして豚の目の解剖という手段があります。その場合、幾分牛と異なる箇所があることを認識しておかなければなりません。
■ 参考 ■
▼以下は「豚の目の解剖」に記述した内容と同じです。
- ヒトの眼は、①網膜に像を結ばせるためのレンズ系,②光の刺激を電気信号に変化させる受容器としての網膜,③生じた活動電位を中枢に伝える神経系の3つに大別される。
- 眼球はほぼ球形をしており、人の眼球の重量は約7.43g,前後径は約24.2mm、周囲は眼窩脂肪体で包まれ保護されている。
- 眼球の壁は3層から成り、外側が強膜という、文字通り強靱な厚い繊維膜になっている。強膜から続く前方約1/6は透明で、この部分を角膜という。
- 強膜の内膜には脈絡膜といわれる血管や色素(メラニン)細胞に富む膜がある。これとつながっている前方は肥厚して毛様体となり、さらにその前方は虹彩となっている。虹彩・毛様体・脈絡膜などを総称してブドウ膜という。
- 眼球の最内側には光の受容を行う網膜となっている。網膜は内側から、①神経繊維層,②神経節細胞層,③内網状層,④内顆粒層,⑤外網状層,⑥外顆粒層,⑦桿体錐体層,⑧色素上皮層(ブルッフ膜)の主な8層からなる。脈絡膜は、内側から(網膜側から)脈絡毛細血管板,血管層,上脈絡膜からなる。
- 物を注視するには、網膜の中心窩(黄斑の中心部で、径が約0.4mmの凹地のこと)に像を結ぶ必要があり、そのために眼球は完全に静止しなくてはならないが、実際には絶えず振動している。この微妙な眼球の振動を生理的眼振(視線固定視眼振)という。これが行われている詳細な理由は明確にされていないが、これによって網膜に巡らされている血管の像が認識できないようになっている。(認識していたとすると視野一面が真っ赤に見えていたり、赤い川が見えていたのかもしれないかもしれませんね。)
- ヒトの瞳孔の直径は2~8mmの範囲で変化する。瞳孔に強い光が入ると0.2~0.5秒の潜時で、虹彩にある瞳孔括約筋が収縮し、瞳孔が縮小する。これを瞳孔の対光反射という。この反射は光の絶対量ではなく、相対的な変化によって生じる。また、この反射は片目だけに光を当てても両眼で生じ、これを共感性光反射と呼ぶ。
- 暗闇の中でフラッシュをたいて写真を撮影したとき、往々にしてヒトの眼は赤く写る。これは強い光が網膜で反射して見える光である。網膜とその後ろの脈絡膜には多くの毛細血管が走っているために、その血管の赤さが光って赤く写る。
- イヌやネコの場合は、網膜に緑色の色素(タペタム)を持っており、緑っぽく写る。ただし、タペタムは網膜全面積の半分程度で、それ以外の半分はヒトと同じであるため色素のない部分で反射するとヒトと同様に赤く写る。タペタムはヒト、ブタ、ラマ、鳥類などには見られない。
- 厚さの割合は、強膜が一番厚く、続いて脈絡膜,網膜の順になる。網膜の厚さはほとんどが神経細胞や支持細胞で、視細胞(桿体、錐体)のある層は、網膜全体の厚さの1割程度である。
- 一般的な眼底写真(--->こちら)で見られる血管は網膜血管で、この血管は網膜の視細胞以外に栄養を供給している。視細胞はその外層の網膜色素上皮から栄養供給されている。脈絡膜の血管は、内層の網膜色素上皮に色素があるために見えません。まれに網膜色素上皮の色素欠損の動物で脈絡膜の血管が見えることがある。この場合眼底が真っ赤に見え、イヌではシベリアンハスキー等に多く見られる。
- 眼は光を受容する感覚器であるが、これは他の動物にも存在し、ミドリムシEuglenaは鞭毛基部の感光点がその働きを担い、正の走光性を示す。光に向かう理由は、ミドリムシの意識ではなく、眼点が一方向からの光を遮って光の方向を探知することによる。
- アメフラシAplysiaの神経節やガンガゼDiademaの放射神経などのニューロンは、光感受性を持つことが知られている。
- プラナリアの杯状眼は、色素層のくぼみの中に視細胞が集まっており、方向視ができる。それがさらに深くくぼんだオウムガイNautilusの眼では形態視が可能である。
- さらに発達すると、節足動物の複眼,軟体動物頭足類および脊椎動物のカメラ眼がある。頭足類と脊椎動物の目は、それぞれ独立に進化したものであるが、よく似た構造を持つ。
- 遠近について、頭足類および魚類・両生類ではレンズと網膜間の距離の変化、爬虫類・鳥類・哺乳類ではレンズの曲率の変化により調節している。
- 頭足類をはじめ、多くの無脊椎動物の目では、視細胞の並ぶ外側(ここでの外側とは、光の来る方向とは逆の方からという意味)から神経繊維が出ている。
- 一方、脊椎動物の網膜では、最外層の色素上皮層の内側にある視細胞から、さらに眼球の内側に向かって神経細胞が層状に配列し、視神経繊維は最内層にある。このような眼を倒立眼という。生物の問題で、“光の来る方向はどちらか”という問題のネタになる。
- 倒立眼の場合、構造上、視神経繊維が視細胞よりも内側にあるため、盲斑(盲点)という視神経の通り道ができ、ここでは視細胞が存在せず、光を受容する能力がない。
- 光を受容する視細胞には、桿体細胞(棒細胞)と錐体細胞(円錐細胞)の2種がある。この名前は先端にそれぞれ桿体状(棒状)と錐体状(円錐状)の突起を持っていることによる。
- ヒトの場合、桿体細胞は中心窩にはなくその周囲に多く分布し、その数は約1億2千万個とされている。錐体細胞は中心窩に集中し、約6千万個あるといわれている。
- 桿体細胞と錐体細胞には外節いわれる円板が積み重なったような膜構造があり、ここに感光物質が存在している。
- 感光物質は光を吸収することにより、その物質の構造が変化し、これによって視細胞のNa+(ナトリウムイオン)透過性が減少し、過分極(受容器電位)が生じる。これが視神経繊維や視神経を介して脳に伝わる。
- 具体的には、桿体細胞内の感光物質であるロドプシン(視紅)に光が当たると、ロドプシン中のシス型レチナールは、トランス型に異性化され、オプシンから遊離される。これにより桿体状外節膜のNa+透過性が低下し、桿体細胞に過分極を起こさせ、電位が生じる。
- 錐体細胞の場合は、吸光性の異なる3種類の感光物質があり、それぞれを含む別々の錐体細胞が存在する(すなわち特定の色に反応する3種類の錐体細胞が存在するということ)。これらの最大吸収波長は、440~450nm(青を吸収する色素:サイアノレーブ),525~535nm(緑を吸収する色素:クロロレーブ),555~565nm(赤を吸収する色素:エリスロレーブ)と言われている。
- 以上の中で抽出されているのは1937年,waldによる、ニワトリの網膜の錐体細胞から抽出された最大吸収波長562nmの紫色の物質であり、ヨドプシンと呼ばれている。このヨドプシンはロドプシンと同様にレチナールとオプシン(錐体細胞のオプシンはフォプシンという)との結合物である。
- 明るい所から暗い所に入って次第に慣れることを暗順応という。暗順応は長時間(30分~1時間)を要し、網膜の光感度は明所での1万倍に達する。一方、明順応は短時間(30秒~1分)で完了する。
- 節足動物の眼には、単眼と複眼がある。複眼は多数の個眼が集まってできている。それぞれの個眼は少しずつ光軸がずれており、複眼全体として視野中の像をモザイクのように合成すると考えられている。
- ミツバチのような昼行性の昆虫では、隣り合う個眼はそれぞれ独立に働く(連立眼)が、蛾のような夜行性のものでは、夜間は個眼の間を仕切る色素粒が移動し、隣接した個眼に入射した光も利用できる(重複眼)。甲殻類の複眼も重複眼である。
- 節足動物の視細胞は、脊椎動物の場合とは異なり、光が当たると脱分極する。
- 昆虫の眼は色覚視ができ、ミツバチの可視部は、ヒトの可視光より短波長側(紫外線側)にずれている。また、ミツバチは光の偏光面を感知する能力を持つ。
|
|
|

